Nature Neuroscience:一种有助于tau种子繁殖和神经毒性的蛋白
2023-01-09 brainnew神内神外 brainnew神内神外 发表于安徽省

研究提高了对tau种子性质的理解,强调了识别相互作用因子的重要性,如BSN,它可以作为致病种子的支架或稳定剂,抑制这些相互作用可能成为治疗神经变性疾病的新方法。
Tau 蛋白聚集是阿尔茨海默病和其他 tau 蛋白病的定义性组织病理学特征。然而,tau 传播所涉及的细胞机制仍不清楚。有研究检测了人类细胞系和诱导的多能干细胞衍生神经元中总 tau 蛋白的相互作用, 但没有研究直接比较具有播种能力的 tau 的相互作用组与单体 tau 的相互作用组,或确定 tau 种子相互作用体如何影响这种种子的性质以及其tau繁殖。
近期,《nature neuroscience》期刊发表了题为“Bassoon contributes to tau-seed propagation and neurotoxicity”的文章,作者团队揭示了一种蛋白,BSN, 如何在体外和体内加剧 tau 的播种和毒性,并证明BSN 下调减少了 tau 蛋白扩散并挽救了 tau 病小鼠模型中的突触结构、功能和行为障碍。 研究结果强调了识别 tau 相互作用因子(例如 BSN)的重要性,它们可以作为 tau 种子的稳定剂,增强其繁殖和毒性。
 1. BSN 蛋白与高分子量 tau 种子相互作用
1. BSN 蛋白与高分子量 tau 种子相互作用
作者从PS19小鼠模型中鉴定了tau种子,该模型过度表达携带p.Pro301Ser(P301S)突变的人tau。为了评估参与播种活动的tau物种的分子量,对3个月大的PS19小鼠的Tris-Buffered生理盐水(TBS)可溶性脑提取物进行了大小排除层析(SEC),这代表了病理学的早期阶段。通过将其导入tau RD P301S荧光共振能量转移(FRET)生物传感器细胞,并用流式细胞仪(图1a)定量测定整合的FRET密度,评估了每个SEC组分的tau种子活性。发现组分9中含有大于2,000 kDa的HMW蛋白质,并具有最强的种子活性(图1b)。
通过对人tau的酶联免疫吸附试验(ELISA)测定,组分9只含有总tau的5%(图1c)。有趣的是,即使在用变性剂盐酸胍(GDN-HCl)处理后,组分9中也含有低百分比的总tau,这揭示了隐藏的表位,意味着在组分9中检测到的低总tau水平并不是由于HMW tau复合体中的表位掩蔽。在野生型(WT) 同窝仔的任何SEC组分中都没有观察到播种活动(图1b)。
为了确认组分9中存在的tau物种是导致播种活性的原因,使用HT7抗体通过免疫沉淀(IP)耗尽了人tau,并观察到流过的tau的播种活性急剧下降(图1d)。电子显微镜(EM)对来自组分9的tau IP材料进行了分析,结果表明,种子以短丝状结构为主,宽度为7±3 nm(图1e)。
使用HT7抗体对来自组分9和组分17(包含单体tau)的人tau进行IP,以识别从同一大脑分离的HMW tau种子和单体tau相互作用的蛋白质。IP产物通过基于TMT标签的定量质谱分析(图1f,g),DAVID 功能注释聚类揭示了两个相互作用组中突触术语丰富, 有趣的是,许多被确认为tau种子相互作用的突触蛋白与那些与单体tau相互作用的突触蛋白不同。我们在“Wall of Targets”中将这两种相互作用组与 AMP-AD 指定的 AD 潜在治疗靶点进行了比较。通过这一比较,作者发现BSN是突触前活动区的支架蛋白,参与调节突触前神经递质释放、突触前蛋白稳定和自噬,是tau种子的重要相互作用因子。令人惊讶的是,最近在一个病理性聚集的3R/4R tau家族中发现了BSN基因的错义突变。此外,BSN在多系统萎缩患者中表达增加,在多发性硬化症患者中积聚。
作者通过免疫共沉淀 (co-IP) 在 PS19 小鼠的 SEC 组分 9 中证实了 BSN 和 tau 种子之间的相互作用(图 1h)。双重染色还揭示了 PS19 小鼠中 BSN 与 tau 沉积物的强烈共定位(图 1i)。BSN 和磷酸化 tau 之间的共定位程度随着年龄的增长而增加。在 tau 病理学的后期阶段(9个月大),两种信号的共定位似乎是在细胞质,但在早期阶段(3 个月大),共定位表现为弥漫性斑点状。考虑到BSN主要是一种突触前蛋白,并且在病理的早期阶段已经在突触前检测到tau,在3个月时对BSN、磷酸化tau PHF1和突触前标记突触蛋白1(Syn-1)进行了三重染色,以表明BSN和Pho-tau共存于突触前终末。这些结果表明,在病理的早期阶段,tau种子与BSN在突触前区相互作用,随着疾病的进展,tau聚集体和BSN共同沉积在细胞质中。
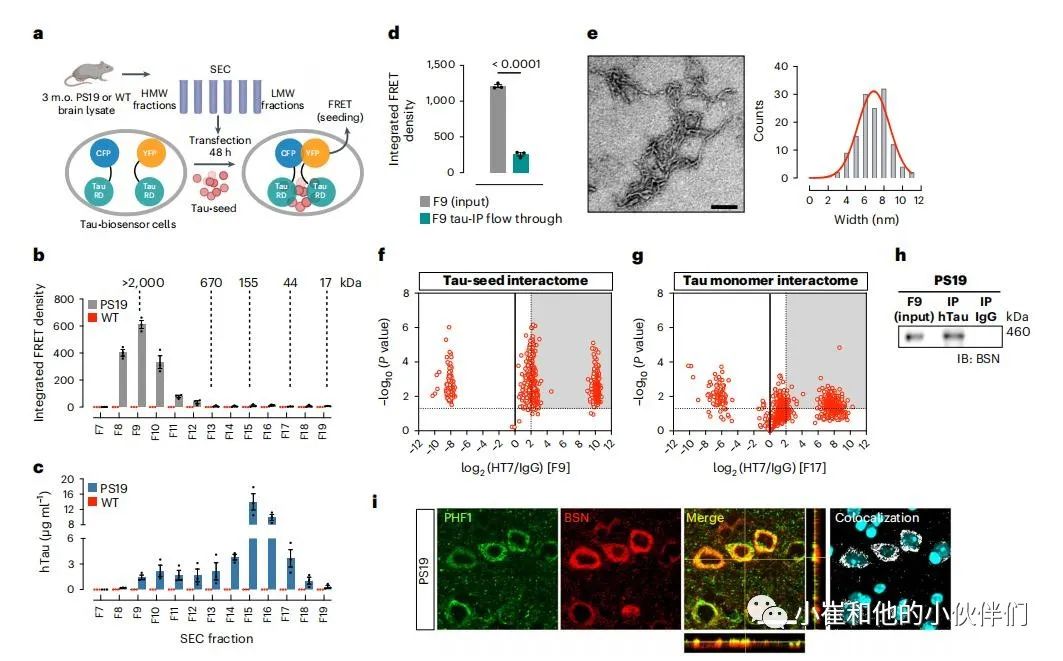
图1:高分子量的tau种子与BSN蛋白相互作用
2. BSN 与人类 tau 病变脑组织中的 tau 病理相关
在AD和PSP患者脑中,种子活性最强的tau物种也是组分9(>2,000 kDa;图2a,b)中的HMW tau物种。与PS19模型中一样,对于两种人类tau病,组分9中的tau只占大脑中总tau的一小部分(图2c,d)。有趣的是,年龄匹配的对照组在组分9中有类似的tau水平,但缺乏种子活性(图2a-d)。从AD和PSP脑中耗尽组分9中的tau显著降低了流经过程中的种子活动(图2e)。组分9的tau IP材料的EM表明,在AD和PSP中,种子以扭曲的细丝为主,宽度分别为6.95±1.1 nm和6.77±1.4 nm(图2f,g)。当从年龄匹配的对照组免疫沉淀tau时,没有检测到tau细丝(图2f)。然后,通过co-IP证实,在这两种tau病中,BSN与组分9中的tau相互作用,但不与年龄匹配对照的组分9中的tau相互作用(图2h)。双重染色显示BSN和tau沉积在AD和PSP脑内共存(图2i)。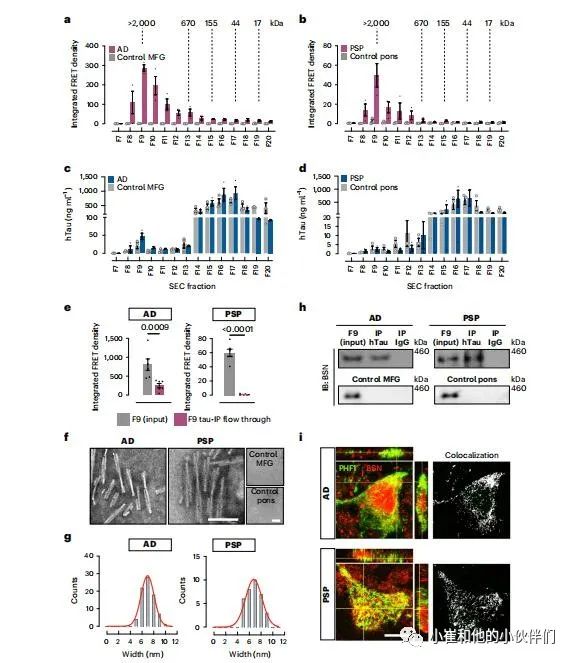
图2:BSN与人类AD和PSP的tau病理有关
3. BSN 增强了tau种子活性和毒性
在人胚胎肾(HEK)293T细胞中过表达人P301S tau蛋白,观察了BSN对tau蛋白病理性质的影响。与只表达P301S tau的细胞的裂解物相比,表达P301S tau和BSN的细胞裂解物的tau种子活性增加(图3a)。BSN的过度表达也与错误折叠的tau物种积累增加有关(图3b)。共表达BSN和P301Stau的HEK细胞双重染色表明,BSN与错误折叠的tau物种共沉积(图3c)。在同一个共表达实验中,通过免疫共沉淀证实了BSN与P301Stau的相互作用(图3d)。在HEK细胞上进行了PLA邻位连接技术以确定BSN是否直接与tau相互作用。共表达P301S tau和BSN的细胞显示出强烈的PLA信号,表明P301S tau和BSN处于相互作用的作用范围内 (<40 nm;图3e)。
考虑到BSN与来自PS19小鼠的tau种子而不是单体tau相互作用(图1f,g),以及与来自AD和PSP患者的组分9中的tau,但不与来自没有种子活性的年龄匹配的对照组的tau相互作用(图2h)。作者的目的是确定这种相互作用是否依赖于构象。由于过表达类似数量的人野生型tau不能形成稳定的聚集体,也没有显示出与人P301S tau相同的种子活性。因此在共表达BSN和人WT tau的HEK细胞上进行了PLA,以确定BSN是否与非聚集性tau相互作用。在过度表达WT tau和BSN的细胞中观察到最小的PLA信号(图3e),表明当tau采用错误折叠或聚集构象时,BSN与tau具有更高的亲和力。
在包括免疫沉淀和质谱仪(IP-MS;图1f)的tau种子分析中,确定了与BSN蛋白的N-末端和C-末端区域相对应的BSN多肽,表明tau种子与全长BSN相互作用。在HEK细胞中与人BSN的N-末端片段(1-850)或C-末端片段(2,450-3,942)共表达P301S tau,并与6x-His标签融合,两个片段在相似的水平上表达,以确定BSN与聚集的tau相互作用的区域。然后,进行了PLA实验,观察到共转染P301S tau和BSN C-末端片段的细胞具有强烈的PLA信号(图3f),表明BSN可以通过其C-末端区域与tau聚集体相互作用。然而,尽管显著降低,但在共转染P301S tau和BSN N-末端片段(图3f)的HEK细胞中观察到了PLA信号,这表明BSN也可以通过其N-末端区域与tau聚集体相互作用。有趣的是,研究表明,疏水性低于平均水平、内在无序残基更多、无序区域更长的大蛋白质更容易与淀粉样聚集体发生异常相互作用。考虑到BSN是一种几乎完全无序的大型蛋白质,并且具有低于平均水平的疏水性,BSN有可能与聚集形式的tau发生广泛的异常相互作用。
在两个转基因黑腹果蝇中过表达人BSN,以研究BSN在体内上调的影响,一个过表达WT BSN,另一个过表达BSN具有p.Pro3866Ala突变,最近在3R/4R tau病理性聚集的患者中被发现。这两个品系都没有发展出退化的眼睛表型(图3g)。然而,WT BSN的过表达通过增加小眼结构的破坏而增强了含有p.pro301leu突变的表达人tau的果蝇模型中的退化眼型(hTau-P301L)(图3g,h)。当BSN突变体过表达时,退行性眼部表型在hTau-P301L中的表达增强(图3g,h)。通过co-IP证实了WT和突变的BSN在果蝇中也与tau相互作用。WB表明,WT和突变体BSN都促进了错误折叠的tau物种的积累,这是由MC1抗体检测到的。然后,使用了胍的稳定性测试来进一步检查果蝇hTau-P301L聚集体之间的稳定性差异。在WT和突变BSN的存在下,tau聚集体明显更耐Gdn-HCl的解聚。正如在细胞模型中观察到的那样,在hTau-P301L果蝇中过表达BSN导致了tau种子活性的增加,在突变的BSN中甚至更高(图3i)。
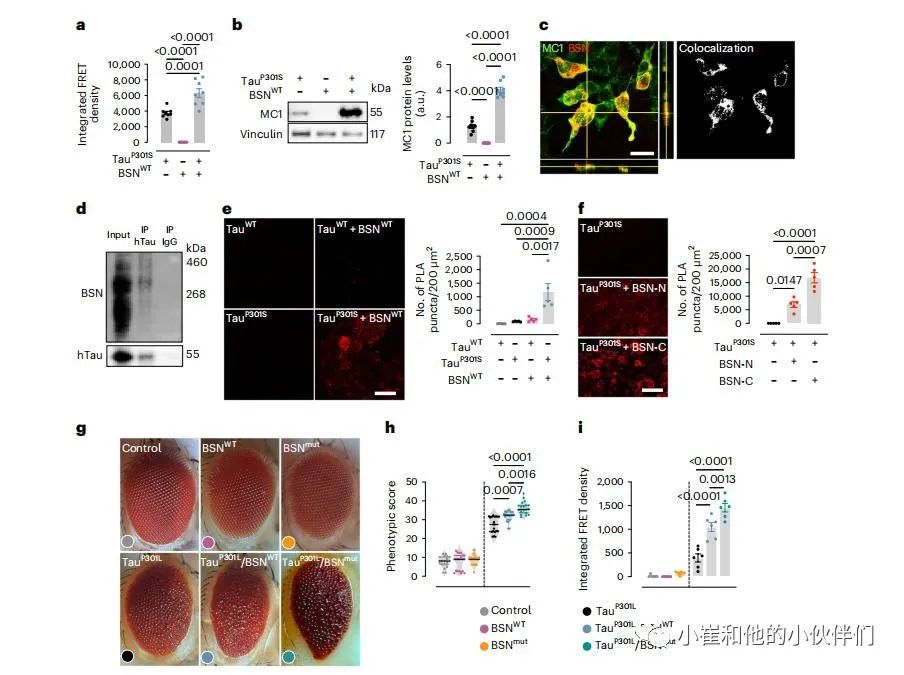
图3:BSN的过度表达通过直接相互作用增加了tau种子活性和毒性
4. BSN基因敲除减少了tau在体内的扩散
使用一个基于腺相关病毒(AAV)的传播模型来研究BSN是否对tau在大脑中的传播起关键作用。在这个模型中,GFP和hTauP301L蛋白是在巨细胞病毒启动子的控制下从mRNA-GFP-P2A-hTauP301L翻译而来的。由于扩散,这个模型允许区分同时表达GFP和hTauP301L的转导神经元和只表达hTauP301L的转导神经元(图4a)。在新生WT小鼠脑内内注射抗BSN的短发夹状RNA(shRNA)的AAV,以同时下调BSN的表达。shBSN可以显著下调WT小鼠体内BSN的表达(至~60%)。scramble和shBSN shRNAs都编码一个蓝色荧光蛋白(BFP2)报告,允许在整个大脑中可视化AAV转导,而不会产生严重异常或对突触前完整性产生负面影响。
具体来说,新生(P0)小鼠接受了pAAV9-mTagBFP2-U6-mBsn-shRNA或对照pAAV9-mTagBFP2-U6-SCR-shRNA的脑室注射,3个月后,将pAAV-GFP-(P2A)-hTauP301L注射到海马区。注射三个月后,小鼠被安乐死,tau扩散通过使用抗hTau HT7抗体的免疫染色来评估(图4b,c)。通过计数每平方毫米中hTau+/GFP细胞的数量来量化两组中tau的扩散(图4c,d)。在注射了scramble AAV的小鼠中观察到大量的tau传播,而在BSN基因敲除的小鼠中传播显著减少(图4d)。这一下降不是由于转导细胞数量的差异,因为在两组中观察到的GFP阳性细胞数量相当(图4d)。
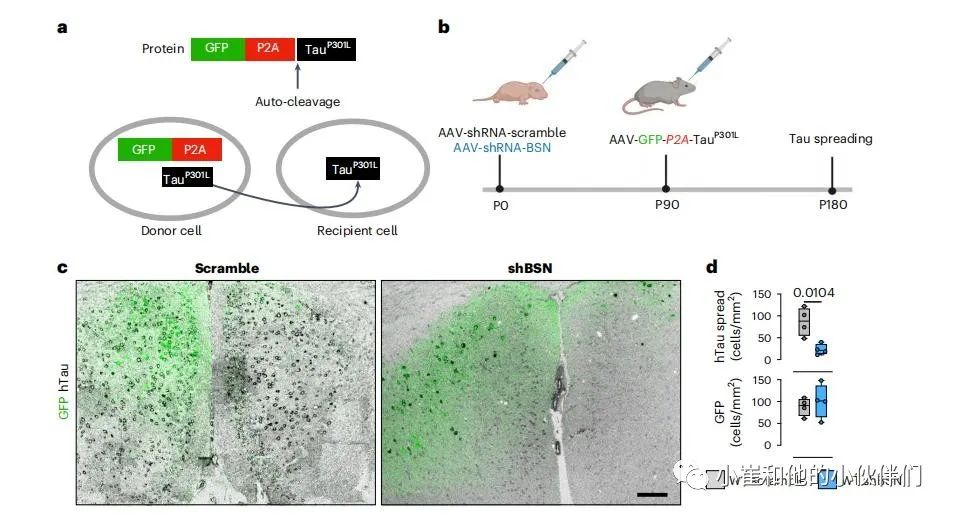
图4:BSN有助于tau在体内的传播
5. BSN下调降低了体内的tau病理和种子稳定性
接下来研究了BSN下调是否影响了PS19小鼠的tau病理和相关的发病机制。给新生PS19和WT仔鼠注射编码scramble或BSN shRNA的AAV。注射4个月后,将小鼠安乐死,并进行WB以确认BSN下调。对抗错误折叠tau抗体(MC1)的检测表明,BSN水平的下降降低了PS19小鼠海马区tau的病理变化(图5a,b)。小胶质细胞和星形胶质细胞同样减少(图5c-e)。在雌性和雄性PS19小鼠中,BSN下调对tau病理和胶质增生的影响都是显著的。在非还原条件下的WB表明,BSN下调降低了总裂解物中的磷酸化tau聚集体(p-tau;使用PHF1(pSer396/Ser404)和pThr231抗体检测到)(图5f-h)。ELISA法显示,BSN下调并不影响PS19脑中人类tau的总水平(图5i);但是,tau种子活性显著下降(图5j)。BSN下调显著降低了雌性和雄性小鼠的PHF1聚集体水平和播种活性,但对pThr231 tau水平的影响仅在雄性PS19小鼠中观察到。然后,对脑裂解物进行了SEC检查,以确定BSN下调是否影响HMW tau种子的活性。正如预期的那样,含有HMW tau种子的组分(组分9)在PS19 shBSN中的种子活性显著低于PS19 scramble样本(图5k)。
有趣的是,在PS19 shBSN和PS19 scramble小鼠的SEC组分中也观察到了类似的总tau水平的分布(图5l),这表明BSN在HMW tau复合体的形成中起到的作用很小,但相反,它可能通过稳定和促进HMW tau种子聚集从而增强其种子活性来影响tau种子的性质。用带有强还原条件的蛋白质印迹分析SEC组分来验证这一假说。在PS19 scramble的样品中,tau在HMW组分(F8、F9和F10)中仍然位于凝胶的顶部,但在PS19 shBSN的样品中,HMW tau以单体的形式在凝胶中溶解(图5m)。当使用MC1抗体时,在PS19 scramble样品中,tau再次保持在HMW SEC组分的凝胶顶部,但在PS19 shBSN样品中,在强还原条件下,没有观察到错误折叠的tau(图5m)。这些数据表明,BSN稳定了HMW tau聚集体,增加了它们在还原条件下的耐降解性。
评估PS19 scramble和PS19 shBSN小鼠脑切片中错误折叠的tau对蛋白酶K(PK)的敏感性,以进一步探索BSN是否影响tau聚集体的结构特性。MC1在PS19 shBSN样本中检测到的错误折叠tau对蛋白酶降解非常敏感,在PK消化45s后强度显著下降。这一结果表明,在没有BSN的情况下,tau聚集体具有松弛的结构,易于消化,而在BSN的存在下,tau聚集体更加致密,对PK具有抵抗力。由于PS19小鼠在3个月大时就发生突触丢失,我们评估了4个月大的PS19小鼠在BSN下调后tau病理的减少是否与突触完整性的改善相关。对突触前(Syn-1)和突触后(PSD95)标记在PS19 scramble和PS19 shBSN小鼠大脑皮质中共定位检测显示(图5n),PS19 shBSN小鼠的突触完整性显著改善(图5o)。BSN下调对PS19小鼠突触完整性的影响可能是由于减少了错误折叠的tau在突触前的积累。
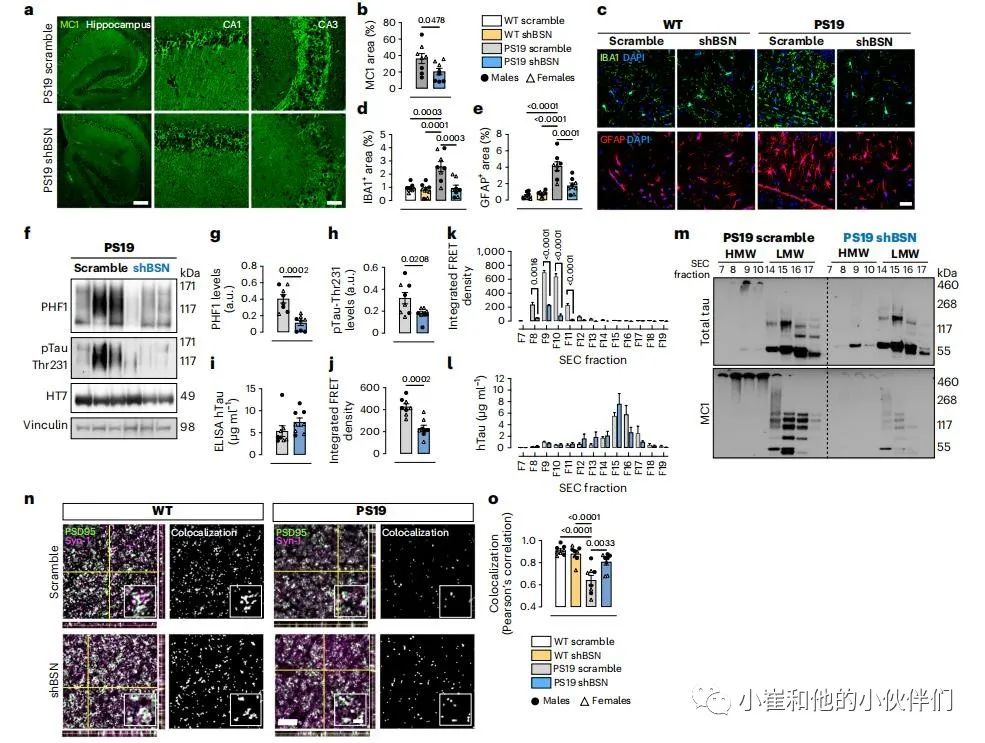
图5:BSN的下调降低了tau的病理和体内tau种子的稳定性
6. BSN降低挽救Tau病小鼠模型的表型
另一组6个月大的PS19和WT仔鼠在P0处注射了scramle或BSN shRNA,以确定这种突触完整性挽救是否代表功能改善。测量了小鼠脑片CA1海马区的长时程增强(LTP)。监测了细胞外刺激Schaffer侧支诱发的场兴奋性突触后电位(FEPSP)通路并采用高频刺激(四个100赫兹刺激1s,间隔10s)诱导LTP。与WT scramble和WT shBSN小鼠相比,PS19 scramble小鼠的海马LTP严重受损;然而,降低BSN水平可以拯救PS19 shBSN小鼠的LTP (图6a-c)。PS19 shBSN小鼠与WT scramble或WT shBSN对照组小鼠的LTP没有差异。还测量了PS19 scramble小鼠与WT scramble和WT shBSN动物相比的配对脉搏比(PPR),反映了突触囊泡释放概率的减少。这种减少在PS19 shBSN动物中得到了挽救(图6d)。在雌性和雄性PS19小鼠中都观察到BSN下调对LTP和PPR的有益影响。
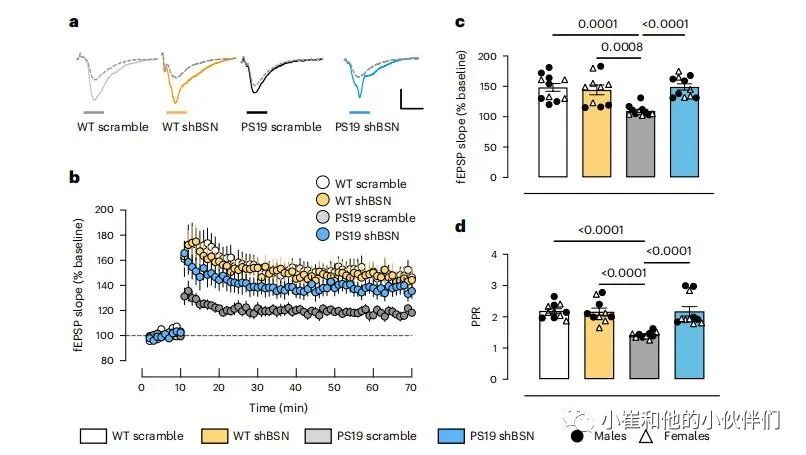
图6:BSN下调可恢复PS19小鼠的电生理损伤
第三组9个月大的小鼠接受了行为和生理测试,以进一步评估BSN下调的功能益处。由于最近对前两只爪子和所有四只爪子的握力进行了测试,已经证明了PS19小鼠运动强度的下降。与WT scramble和WT shBSN小鼠相比,PS19scramble小鼠的两爪和四爪握力都有所降低(图7a、b)。PS19 shBSN小鼠与WT scramble或WT shBSN小鼠之间无差异,提示BSN下调可挽救运动损伤。通过测定基础核心体温和虚弱指标来评价其生理特性。PS19 scramble小鼠的基础体温低于WT scramble和WT shBSN小鼠;然而,BSN水平的下调完全挽救了PS19 shBSN小鼠降低的体温(图7c)。
然后,进行了临床检查,以评估26个虚弱参数。PS19 scramble小鼠比WT scramble和WT shBSN小鼠表现出更多的脆弱标记(图7d),表明一般健康状况下降。在PS19 shBsn小鼠和任一WT组之间没有观察到差异,这表明在tau病理的背景下BSN下调是有益的。在雄性和雌性PS19小鼠中都观察到了BSN下调可以挽救PS19小鼠的运动和生理。
接下来测试了BSN下调是否可以减少PS19小鼠的神经退行性变。9个月龄的PS19小鼠当脑室容量的增加和海马区体积的减少时表明脑萎缩,通过体积磁共振成像MRI分析,确定了BSN下调对脑室和海马区体积的影响。BSN下调并没有显著改变WT小鼠的脑室或海马体体积。与WT scramble和WT shBSN小鼠相比,PS19 scramble小鼠的脑室体积增加,而海马体体积减少。然而,与PS19小鼠相比,降低PS19小鼠的BSN水平显著改善了心室容量的增加。PS19 shBSN小鼠与WT scramble或WT shBSN小鼠之间的海马体体积没有差异, 提示BSN下调在一定程度上减少神经退行性变。由于小鼠数量有限,不可能确定BSN下调对恢复脑室和海马体体积的有益影响的性别差异。
最后,评估了在9个月大的PS19小鼠中观察到的脑萎缩的功能改善和挽救是否与病理tau负担的减少有关。免疫荧光分析显示,BSN下调降低了PS19小鼠海马区的tau病理(图7e,f)。利用WB,还证实了BSN的下调减少了磷酸化的tau聚集体(图7g-i),但不影响用ELISA法在9月龄PS19小鼠的大脑样本中测量的总tau水平(图7j)。
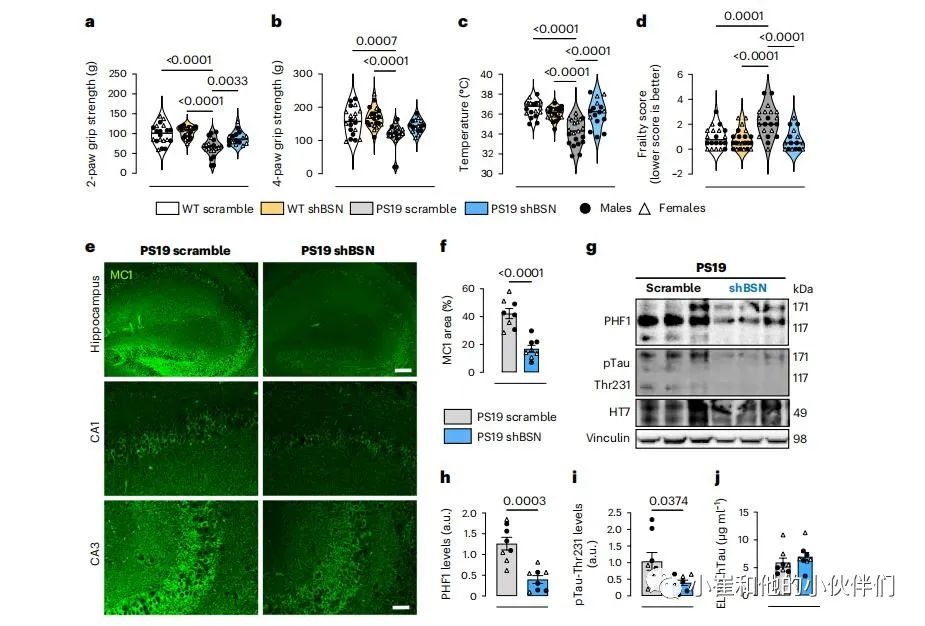
图7:降低BSN水平可改善行为缺陷,并减少病理晚期的病理性tau
小 结
作者团队的研究提高了对tau种子性质的理解,强调了识别相互作用因子的重要性,如BSN,它可以作为致病种子的支架或稳定剂,抑制这些相互作用可能成为治疗神经变性疾病的新方法。
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言



